Journal of Infection and Molecular Biology
Review Article
Journal of Infection and Molecular Biology 2 (2): 26 – 29The Structure and Function of Jembrana Disease Virus Genome
Asmarani Kusumawati1,2*, Tenri Ashari Wanahari3, Rizqa Febriliany Putri3, Basofi Ashari Mappakaya3, Issabellina Dwades Tampubolon2
- Departement of Reproduction, Faculty of Veterinary Medicine, Gadjah Mada University, Jalan sekip unit II, Yogyakarta, 55281 Indonesia
- Centre of Biotechnology Study, Gadjah Mada University, Jalan teknika utara, Yogyakarta, 55281 Indonesia
- Faculty of Medicine, Sebelas Maret University, Jalan Ir. Sutami 36 A, Surakarta, 57126 Indonesia
*Corresponding author: kartapati_2008@yahoo.com
ARTICLE CITATION:
Kusumawati A, Wanahari TA, Putri RF, Mappakaya BA, Tampubolon ID (2014). The structure and function of Jembrana disease virus genome. J. Inf. Mol. Biol. Sci. 2 (2): 26 – 29.
Received: 2014–02–20, Revised: 2014–03–28, Accepted: 2014–03–28
The electronic version of this article is the complete one and can be found online at
(
http://dx.doi.org/10.14737/jimb.2307-5465/2.2.26.29
)
which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited
ABSTRACT
The pathogenic agent of Jembrana disease is a virus which was consequently named Jembrana disease virus (JDV). JDV was definitively defined as a member of the subfamily of lentiviruses of the family of Retroviridae. The genome of JDV is a linear single–stranded RNA, 7732 nucleotides in length. It contains the structural genes gag, pol and env which encode structural and enzymatic proteins that make up infectious viral particles and flanking long terminal repeats (LTR) that characterize all retroviruses. Accessory genes of JDV include tat, rev and vif that are typical of lentiviruses and which are required for the production of regulatory proteins. Products of most accessory genes have been shown to play important roles in viral replication and pathogenesis. The genome of BIV, another bovine lentivirus, is the most related lentivirus to JDV. Though the two bovine lentiviruses have to be considered as two distinct entities as the two viruses are sufficiently different.
INTRODUCTION
Jembrana disease is a bovine disease that affects Bali cattle (Bos javanicus). It was first recognized in 1964 as an acute and infectious disease in the Jembrana District of Bali Island in Indonesia (Budiarso, Hardjosworo, 1976). Due to this report, the disease was therefore designated Jembrana disease. The evidence for a viral aetiology of Jembrana disease was established in 1992. The pathological agent is a lentivirus, consequently called Jembrana disease virus (JDV) (Wilcox et al., 1992). The disease is now endemic through–out parts of Indonesia, including Java, Sumatra (Hartaningsih et al., 1993), and more recently Kalimantan (Borneo), and also in Australia (Chadwick et al., 1998). Jembrana disease constitutes the main concern in Bali cattle industry as it has caused important economic losses due to mortality of cattle. Indeed, during the first observed outbreaks, in 12 months, about 60000 animals died consequently to JDV infection.
The viral nature of the pathogenic agent of Jembrana disease was firstly identified by its size, determined by membrane filtration. It was further established as a member of the family Retroviridae, due to the associated disease, the reverse transcription (RT) activity, the morphological characteristics, the virus maturation by C–type budding through the plasma membrane and into the intracytoplasmic vacuoles, formation of spherical virus particles of 96–124 nm in diameter with smooth membrane which is characteristic of enveloped viruses and frequently with an eccentric nucleoid observed under electron microscopy (Kertayadnya et al., 1993; Wilcox et al., 1992). Furthermore, Western immunoblotting using sera from recovered animals identified several proteins with sizes ranging from 14 kDa to 100 kDa. Among these antigens, a 24 kDa protein cross–reacted with the 26 kDa capsid protein of bovine immunodeficiency virus (BIV) of the lentivirus subfamily. This analysis allowed to definitively affiliate JDV to the subfamily lentiviruses of the family Retroviridae (Kertayadnya et al., 1993; Wilcox et al., 1995).
The lentivirus affiliation was established by comparison of pol gene sequences (Chadwick et al., 1995a). The subfamily of lentiviruses of the Retroviridae family includes many viruses that cause important diseases such as human immunodeficiency virus type 1 (HIV–1) and type 2 (HIV–2), simian immunodeficiency virus (SIV), equine infectious anaemia virus (EIAV), feline immunodeficiency virus (FIV), caprine arthritis encephalitis virus (CAEV), ovine maedi–visna virus, in addition to JDV and BIV.
Genome of JDV
The nucleotide sequence of the JDV genome and the deduced amino acid sequence have now been entirely established. The genome of JDV is a linear single–stranded RNA (GenBank acc. no. NC_001654; Chadwick et al., 1995b). The genome is 7732 nucleotides in length. Due to this size, it is the smallest lentivirus genome up to date reported. Sequence alignment has also enabled the identification of putative features in JDV genome. It contains the structural genes gag, pol and env which encode structural and enzymatic proteins that make up infectious viral particles and flanking long terminal repeats (LTR) that characterize all retroviruses and finally a number of accessory genes, including tat, rev and vif that are typical of lentiviruses and which are required for the production of regulatory proteins (Chadwick et al., 1995b). Products of most accessory genes have been shown to play important roles in viral replication and pathogenesis (Kirchhoff et al., 1999).
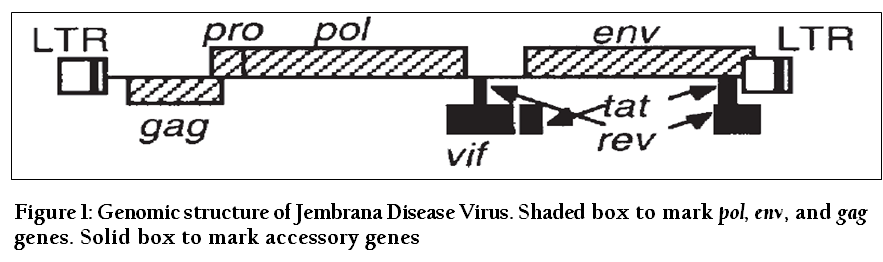
Figure 1: Genomic structure of Jembrana Disease Virus. Shaded box to mark pol, env, and gag genes. Solid box to mark accessory genes
Long Terminal Repeats (LTR)
The presence of long terminal repeats (LTRs), flanking both ends of the genome, is one of the characteristics of retroviral genome. The putative JDV LTR is 397 nucleotides long, delineated by the 17 nucleotides primer binding site at the 3’–end and a 18 nucleotides poly-purine tract at the 5’–end. Other features identified were transcription termination signals and 7 nucleotides inverted repeats found at the junction of the chromosomal DNA and the LTRs. Like in other retroviral genomes, JDV LTRs include three regions, successively U3, R and U5. The U3 region, 236 nucleotides long (148 nucleotides shorter than BIV U3), contains enhancer and promoter elements as well as the TATA box which is thought to be essential to the eukaryotic basal promoter but the CAT box, found in most eukaryotic and retroviral promoters is lacking. The downstream U3 region is the putative R region which is 108 nucleotides long and 3 nucleotides shorter than the BIV R region. Within this region, the poly (A) signal and the termination transcription signal are conserved with regards to the BIV ones. The putative 5’–terminus of the viral RNA genome has been identified as the trans–activating response (TAR) region (Chadwick et al., 1995b) to which the trans–activator of transcription (Tat) protein binds for its activity (Carpenter et al., 1992). JDV TAR region has the potential to form a secondary structure, i.e. stem–loop structures, which is more complex than the BIV one. The most downstream region is the U5 region which is 53 nucleotides long and 42 nucleotides shorter than BIV U5 region. The terminal 7 nucleotides repeat (see above) and the tetranucleotide sequence which is supposed to be implicated with transcription termination, are found both in BIV and JDV genomes (Chadwick et al., 1995b).
Structural Genes
Gag Gene
Gag gene is one of the three structural genes, gag, pol and env (Chadwick et al., 1995b). The gag gene is translated as a single precursor protein which is then cleaved by proteases into three components, i.e. matrix protein (MA) (located at the N–terminal part), capsid protein (CA) which is also the immunodominant virion protein in infected cattle and nucleocapsid protein (NC) (localized at the C–terminal part). The putative JDV gag precursor (Pr53gag) which is 436 amino acids in length, with a calculated molecular weight of ca. 49 kDa, contains two cleavage sites, instead of five in BIV. The processing of JDV Pr53gag should give rise to (i) MA of 125 amino acids, with a molecular weight estimated to be ca. 14 kDa, (ii) CA (or Cp26), composed of 226 amino acids, with molecular weight of ca. 26 kDa, and NC, 85 amino acids in length and a ca. 9 kDa molecular weight. The three additional BIV short processed products p2L (of 22 amino acids, with a molecular weight of 2.5 kDa), p3 (of 25 amino acids, with a molecular weight of 2.7 kDa) and p2 (of 18 amino acids, with a molecular weight of 1.9 kDa) should be missing in JDV (Chadwick et al., 1995b; Lu et al., 2002). In fact, p2L would be actually missing or incorporated into CA, except for the first three N–terminal amino acids (Lu et al., 2002) and p3 and p2 would be incorporated into NC. At the amino acid level, there is a 62% similarity in gag gene products whereas a 75% similarity is found in the capsid protein between JDV and BIV. This strong sequence conservation can so explain the observed serological cross–reactivity between the capsid proteins of the two lentiviruses (Kertayadnya et al., 1993).
Gag–pol Gene
Like in most retroviruses and all lentiviruses, JDV Pol ORF is translated as a gag–pol poly-protein by a –1 ribosomal frame shift. The putative hepta-nucleotide involved in frame shifting mechanism, has also been identified. Pol ORF encodes a protease (PR), a reverse transcriptase (RT) and an integrase (IN) proteins (Chadwick et al., 1995b). The predicted gag–pol precursor protein of JDV is composed of 1432 amino acids with an estimated molecular weight of 163 kDa. The translation requires a frameshift at nucleotide 1444. The pol protein precursor which is translated from the pol ORF itself, is 1027 amino acids long and exhibiting a calculated molecular weight of 118 kDa. The ORFs coding for PR, RT, and IN have not yet been determined. The similarity of the gag gene is approximately 62% at the amino acid level between JDV and BIV.
Env Gene
The product of env gene is expressed as a single spliced mRNA. It is translated into an env precursor of 781 amino acids in length with an estimated molecular weight of ca. 88 kDa. This precursor is further cleaved at a R–A motif into a surface protein (SU), of 422 amino acids in length and a calculated molecular weight of ca. 47 kDa and a transmembrane protein (TM) of 359 amino acids and a calculated molecular weight of ca. 41 kDa (Kusumawati et al., 2010; Chadwick et al., 1995b). A 31% amino acid identity was found between JDV and BIV env gene products overall, with 46% substitutions and 23% mismatches associated with deletions and insertions. JDV TM has a similar size to BIV TM whereas due to a deletion of 471 nucleotides in the SU region, JDV SU is substantially smaller than its BIV counterpart.
Accessory Genes
JDV genome possesses in the central and 3’–terminal domains at least three accessory genes typical of lentivirus genomes, i.e. vif (viral infectivity factor), tat (trans–activator of transcription), and rev (regulator of virus expression). In HIV–1, products of most accessory genes are thought to be functionally implicated in viral replication and pathogenesis. Among them the trans–activator of transcription (Tat) protein which is encoded by tat gene, is probably the most important element required for virus life cycle (Arya et al., 1985; Dayton et al., 1986;
Huang et al., 1994; Kirchhoff et al., 1999; Michael et al., 1995; Westendorp et al., 1995). In JDV, the tat gene and its product, i.e. JDV Tat protein (jTat), are the most thouroughly studied among the accessory genes and the proteins they encode. jTat is thought to also play essential functions in virus life cycle and pathogenesis. Furthermore, this protein is a potent trans–activator of transcription, able to activate its own LTR as well as the LTR of other lentiviruses (Calabro et al., 2005; Chen et al 1999b; 2000; Smith et al., 2000; Su et al., 2009; Xie et al., 2004). Its ability to be internalized in noninfected cells and to interfere with cellular gene expression and with cellular cytoskeleton may constitute one of the keys of JDV pathogenesis (Deng et al., 2006; Xuan et al., 2007).
Tat Gene
The JDV tat gene includes two distanly located open reading frames (ORFs), i.e. coding exon 1 and coding exon 2. The putative jTat protein (jTat), deduced from the nucleotide sequence, is composed of 97 amino acids with a calculated molecular weight of ca. 11 kDa or alternatively of 114 amino acids in length with a calculated molecular weight of ca. 13 kDa. jTat contains a highly conserved cysteine–rich region which presents a 77% identity with that of BIV Tat (bTat). However there is only 8% of sequence identity at the C–terminal domain between jTat and bTat (Chadwick et al., 1995b). Other domains of the 97 amino acids jTat are N–terminal domain, upstream the cysteine–rich region, then successively core domain, basic domain and C–terminal domain (Su et al., 2009).
Like in other lentiviruses, jTat is thought to be produced from multiply spliced mRNAs, due to the presence of numerous splice donors (SD) and splice acceptors (SA), (Chadwick et al., 1995b; Setiyaningsih et al., 2008; Stewart et al., 2008). The translation of exon–1, with an ORF of 294 nucleotides in length, is thought to be sufficient to generate a functional jTat (Chen et al 1999b; 2000; Smith et al., 2000; Su et al., 2009) as for e.g. bTat (Liu, et al., 1992). This JDV tat ORF will give rise to a deduced jTat of 97 amino acid residues with a calculated molecular weight of ca.11 kDa. The apparent molecular weight of native Tat protein, established by Western blot analysis, is 14 kDa (Setiyaningsih et al., 2008). This discrepancy may be due to the presence of positively charged basic residues such as arginine and lysine (Hames, 1998) or post–translational modifications.
Rev Gene
The putative rev protein is also produced from two coding exons, separately located within the genome. They would generate a 213 amino acids protein, with a calculated molecular weight of ca. 24 kDa (Chadwick et al., 1995b) which proves to be 27 amino acids longer than BIV Rev Protein (Oberste et al., 1993). By using the alternative SD in exon 1, the translation of rev gene will result in the production of a 201 amino acids protein, with a calculated molecular weight of ca. 23 kDa. As in most lentiviruses, the greater part of Rev Protein is produced from exon 2. Only 35% amino acid sequence identity is found between JDV and BIV Rev Proteins. Nothing is known, for the time being, on the role of JDV rev protein in viral pathogenesis or in other functions.
Vif Gene
The putative JDV vif gene found in JDV genome has a single potential coding sequence (CDS). Its product is a 197 amino acids protein, with a calculated molecular weight of ca. 23 kDa. Up to now, the function of JDV vif protein is not known.
The Comparison of Genome of JDV and BIV
BIV is the most related lentivirus to JDV. The genome of BIV is a linear single–stranded RNA of 8960 nucleotides in length (GenBank acc. no. M32690 [BIV127]; Garvey et al., 1990). The comparison of genome of BIV and JDV has verified some differences between the two genomes (Chadwick et al., 1995b). The JDV genome is much shorter and strikingly many deletions and insertions have been found relative to that of BIV. Despite the error–prone nature of lentiviral reverse transcriptases (RT) and the genomic variability in vivo, it has never been observed before that two closely related lentiviruses feature so many deletions and insertions rather than substitutions relative to each other. Despite being 750 nucleotides shorter, the genomic organization of JDV genome is almost identical to that of BIV. A high degree of nucleotide and amino acid homology can be observed throughout the two genomes. The presence of small open reading frames (ORFs) in central and 3’–terminal domains also shows that the JDV genomic organization is typical to lentiviruses. Relative to BIV genome, JDV genome shows 27 deletions, totalling 897 nucleotides, and 17 insertions, totalling 147 nucleotides, and resulting so in 750 nucleotides shorter genome (Chadwick et al., 1995b). Comparison of 9 partial pol (polymerase) gene sequences from known lentiviruses has shown that the closest alignment of JDV pol gene sequence is found with that of BIV, indicating a 74% identity at the nucleotide level and 76% at the level of amino acid, and importantly with no deletions or insertions. This analysis definitively confirms that JDV is actually a lentivirus. On the basis of partial pol gene sequence, JDV is the most related to but distinguishable from BIV. The phylogenetic analysis also estimates that the genetic distance between JDV and BIV is comparatively shorter than that between the two human immunodeficiency viruses, HIV–1 and HIV–2 (Chadwick et al., 1995a).
Concluding Remarks
Although BIV, another bovine lentivirus, is genetically and antigenically the most related to JDV, the two bovine lentiviruses have to be considered as two distinct entities as the two viruses are sufficiently diverging, due to important deletions and insertions (Chadwick et al., 1995b). Another reason to consider the two viruses as separate entities is the fact that the associated disease syndromes radically differ each from the other. In contrast to the mild disease syndrome associated with most lentiviruses, including BIV, JDV infection is associated with an acute, severe and often lethal disease. This difference may wholly or partially result from differences found in their genome and may explain the markedly increased level of viral expression and in vivo pathogenicity of JDV. This may result from more efficient enhancer elements, changes in rev gene products or a much more potent Tat protein.
ACKNOWLEDGEMENT
This work was partly funded by a grant from Directorate General of Higher Education (DIKTI), Ministry of Education and Culture of Indonesia.
REFERENCES
Arya SK, Guo C, Josephs SF, Wong–Staal F (1985). Trans–activator gene of human T–lymphotropic virus type III (HTLV–III). Sci. 229: 69 – 73.
http://dx.doi.org/10.1126/science.2990040
Budiarso IT, Hardjosworo S (1976). Jembrana disease in Bali cattle. Aust. Vet. J. 52: 97.
http://dx.doi.org/10.1111/j.1751-0813.1976.tb13867.x
PMid:990093
Calabro V, Daugherty MD, Frankel AD (2005). A single intermolecular contact mediates intramolecular stabilization of both RNA and protein. Proc. Natl. Acad. Sci. U S A. 102: 6849 – 6854.
http://dx.doi.org/10.1073/pnas.0409282102
PMid:15857951 PMCid:PMC1100766
Carpenter S, Miller LD, Alexandersen S, Whetstone CA, VanDerMaaten MJ, Viuff B, Wannemuehler Y, Miller JM, Roth JA (1992). Characterization of early pathogenic effects after experimental infection of calves with bovine immunodeficiency–like virus. J. Virol. 66: 1074 – 1083.
PMid:1309889 PMCid:PMC240811
Chadwick BJ, Coelen RJ, Sammels LM, Kertayadnya G, Wilcox GE (1995a). Genomic sequence analysis identifies Jembrana disease virus as a new bovine lentivirus. J. Gen. Virol. 76: 189 – 192.
http://dx.doi.org/10.1099/0022-1317-76-1-189
PMid:7844531
Chadwick BJ, Coelen RJ, Wilcox GE, Sammels LM, Kertayadnya G (1995b). Nucleotide sequence analysis of Jembrana disease virus: a bovine lentivirus associated with an acute disease syndrome. J. Gen. Virol. 76: 1637 – 1650.
http://dx.doi.org/10.1099/0022-1317-76-7-1637
PMid:9049370
Chadwick BJ, Desport M, Brownlie J, Wilcox GE, Dharma DM (1998). Detection of Jembrana disease virus in spleen, lymph nodes, bone marrow and other tissues by in situ hybridization of paraffin–embedded sections. J. Gen. Virol. 79: 101 – 106.
PMid:9460930
Chen H, He J, Fong S, Wilcox G, Wood C (2000). Jembrana disease virus Tat can regulate human immunodeficiency virus (HIV) long terminal repeat–directed gene expression and can substitute for HIV Tat in viral replication. J. Virol. 74: 2703 – 2713.
http://dx.doi.org/10.1128/JVI.74.6.2703-2713.2000
PMid:10684286 PMCid:PMC111760
Chen H, Wilcox G, Kertayadnya G, Wood C (1999b). Characterization of the Jembrana disease virus tat gene and the cis– and trans–regulatory elements in its long terminal repeats. J. Virol. 73: 658 – 666.
PMid:9847371 PMCid:PMC103872
Dayton AI, Sodraski JG, Rosen CA, Goh WC, Haseltine WA (1986). The trans–activator gene of the human T cell lymphotropic virus type III is required for replication. Cell. 44 : 941 – 947.
http://dx.doi.org/10.1016/0092-8674(86)90017-6
Deng G, Qiao W, Su Y, Sha R, Geng Y, Chen Q (2006). Internalization of Jembrana disease virus Tat: possible pathway and implication. Virus Res. 121: 122 – 133.
http://dx.doi.org/10.1016/j.virusres.2006.01.023
PMid:16870296
Garvey KJ, Oberste MS, Elser JE, Braun MJ, Gonda MA (1990). Nucleotide sequence and genome organization of biologically active provirus of the bovine immunodeficiency–like virus. Virol. 175: 391 – 409.
http://dx.doi.org/10.1016/0042-6822(90)90424-P
Hames BD (1998). Gel Electrophoresis of proteins: A practical approach. 3rd ed. Oxford University Press, New York.
Hartaningsih N, Wilcox GE, Dharma DM, Soetrisno M (1993). Distribution of Jembrana disease in cattle in Indonesia. Vet. Microbiol. 38: 23 – 29.
http://dx.doi.org/10.1016/0378-1135(93)90072-F
Huang LM, Joshi A, Willey R, Orenstein J, Jeang KT (1994). Human immunodeficiency viruses regulated by alternative trans–activators: genetic evidence for a novel non–transcriptional function of Tat in virion infectivity. EMBO J. 13: 2886 – 2896.
PMid:8026473 PMCid:PMC395170
Kertayadnya G, Wilcox GE, Soeharsono S, Hartaningsih N, Coelen RJ, Cook RD, Collins ME, Brownlie J (1993). Characteristics of a retrovirus associated with Jembrana disease in Bali cattle. J. Gen. Virol. 74: 1765 – 1778.
http://dx.doi.org/10.1099/0022-1317-74-9-1765
PMid:7690840
Kirchhoff F, Easterbrook PJ, Douglas N, Troop M, Greenough TC, Weber J, Carl S, Sullivan JL, Daniels RS (1999). Sequence variations in human immunodeficiency virus type I Nef are associated with different stages of disease. J. Virol. 73: 5497 – 5508.
PMid:10364298 PMCid:PMC112607
Kusumawati A, Pratiwi R, Astuti P, Hamid PH (2010). Characterization of envelope-transmembrane gene of jembrana disease virus tabanan 1995 isolate. Indo. J. Biotechnol. 15: 15 – 19.
Liu ZQ, Sheridan D, Wood C (1992). Identification and characterization of the bovine immunodeficiency–like virus tat gene. J. Virol. 66: 5137 –5140.
PMid:1321293 PMCid:PMC241389
Lu M, Zheng L, Mitchell K, Kapil S, Wood C, Minocha H (2002). Unique epitope of bovine immunodeficiency virus gag protein spans the cleavage site between p16 (MA) and p2L. Clin. Diagn. Lab. Immunol. 9: 1277 – 1281.
PMid:12414761 PMCid:PMC130104
Michael NL, Chang G, d'Arcy LA, Ehrenberg PK, Mariani R, Busch MP, Birx DL, Schwarts DH (1995). Defective accessory genes in a human immunodeficiency virus type 1–infected long–term survivor lacking recoverable virus. J. Virol. 69: 4228 – 4236.
PMid:7769682 PMCid:PMC189160
Oberste MS, Williamson, JC, Greenwood JD, Nagasima K, Copeland TD, Gonda MA (1993). Characterization of bovine immunodeficiency virus rev cDNA and identification and subcellular localizatiuon of the rev protein. J. Virol. 67: 6395 – 6404.
PMid:8411341 PMCid:PMC238074
Olsen JC (1998). Gene transfer vectors derived from equine infectious anemia virus. Gene Ther. 5: 1481 – 1487.
http://dx.doi.org/10.1038/sj.gt.3300768
PMid:9930301
Setiyaningsih S, Desport M, Stewart ME, Hartaningsih N, Wilcox GE (2008). Sequence analysis of mRNA transcripts encoding Jembrana disease virus Tat–1 in vivo. Virus Res. 132: 220 – 225.
http://dx.doi.org/10.1016/j.virusres.2007.11.004
PMid:18096262
Smith CA, Calabro V, Frankel AD (2000). An RNA–binding chameleon. Mol. Cell. 6: 1067 – 1076.
http://dx.doi.org/10.1016/S1097-2765(00)00105-2
Stewart ME, Desport M, Setiyaningsih S, Hartaningsih N, Wilcox GE (2008). Analysis of Jembrana disease virus mRNA transcripts produced during acute infection demonstrates a complex transcription pattern. Virus Res. 135: 336 – 339.
http://dx.doi.org/10.1016/j.virusres.2008.03.017
PMid:18466992
Su Y, Deng G, Gai Y, Li Y, Gao Y, Du J, Geng Y, Chen Q, Qiao W (2009). Comparative functional analysis of Jembrana disease virus Tat protein on lentivirus long terminal repeat promoters: evidence for flexibility at its N–terminus. Virol. J. 6: 179 – 193.
http://dx.doi.org/10.1186/1743-422X-6-179
PMid:19860923 PMCid:PMC2775740
Westendorp MO, Frank R, Ochsenbauer C, Striker K, Dhein J, Walczak KH, Debatin KM, Kramer PH (1995). Sensitization of T cells to CD95-mediated apoptosis by HIV-1 Tat and gp 120. Nature. 375: 497 – 590.
http://dx.doi.org/10.1038/375497a0
PMid:7539892
Wilcox GE, Chadwick BJ, Kertayadnya G (1995). Recent advances in the understanding of Jembrana disease. Vet. Microbiol. 46: 249 – 255.
http://dx.doi.org/10.1016/0378-1135(95)00089-S
Wilcox GE, Kertayadnya G, Hartaningsih N, Dharma DM, Soeharsono S, Robertson T (1992). Evidence for a viral aetiology of Jembrana disease in Bali cattle. Vet. Microbiol. 33 : 367 – 374.
http://dx.doi.org/10.1016/0378-1135(92)90064-Z
Xie B, Calabro V, Wainberg MA, Frankel AD (2004). Selection of TAR RNA–binding chameleon peptides by using a retroviral replication system. J. Virol. 78: 1456 – 1463.
http://dx.doi.org/10.1128/JVI.78.3.1456-1463.2004
PMid:14722301 PMCid:PMC321383
Xuan C, Qiao W, Gao J, Liu M, Zhang X, Cao Y, Chen Q, Geng Y, Zhou J (2007). Regulation of microtubule assembly and stability by the transactivator of transcription protein of Jembrana disease virus. J. Biol. Chem. 282: 28800 – 28806.
http://dx.doi.org/10.1074/jbc.M702823200
PMid:17656359



